Content of Fe and Mn in leaves and stems, and in seeds macronutrients were determined in Dion edule (chamal). Chamal is a palm belonging to cycads, used as an ornamental plant and seeds as a substitute for corn for human consumption in some indigenous communities in Huasteca Potosina called xi'oi. Concentration of Fe and Mn were evaluated in stems, leaves and seeds by atomic absorption spectrophotometry to determine micronutrients relationship in the whole plant and its translocation to seed. Sand moisture content, ash, protein, fat and carbohydrates, was determined only in seeds according to official proximal chemical analysis techniques for food. Concentrations of Fe and Mn were higher in leaves than in stem. Regarding macronutrient content in seeds was found D. edule they contain 14.67% moisture, 20.65% protein, 3.21% lipids and 60.69% carbohydrates. Results show that seeds nutrient content of D. edule is similar to dry-bean and greater than corn, which are a suitable source of Fe and Mn therefore constitute an alternative to prepare nutritious food source.
INTRODUCCIÓN
Las cycadales son consideradas uno de los órdenes de plantas con mayor periodo evolutivo, ya que son las plantas con semilla más antiguas del mundo; se encuentran distribuidas en el continente africano, americano, asiático y oceanía (Brener, Stevenson & Twig, 2003). México es el segundo país con mayor riqueza en especies de cycadales, con aproximadamente un 20% de las especies del mundo, de las cuales el 80% son endémicas. En este país solo se distribuye la familia Zamiaceae, dentro de la cual se encuentran tres géneros Dioon, Zamia y Ceratozamia con 12, 15 y 21 especies, respectivamente (Yáñez, 2006).
En la Huasteca Potosina, ubicada al noreste de México, la distribución de cycadales se acentúa más que en otras regiones del país, donde los habitantes de la etnia xi’oi practican la agricultura basada en el maíz, frijol y caña de azúcar. Además, llevan a cabo la recolección de plantas cycadales como Dioon edule conocido como chamal. La cosecha de D. edule tiene un papel importante en su dieta y la usan como un sustituto del maíz. Entre los alimentos que la etnia xi’oi elaboran a partir de la masa obtenida a base de semillas hervidas y machacadas de D. edule, se incluyen tortillas, gorditas y tamales. Debido al uso de esta semilla para la elaboración de alimentos básicos resulta de interés conocer el contenido de macronutrientes que esta pudiera aportar a la dieta de quienes la consumen. La calidad de los alimentos no está determinada solo por el contenido de macronutrientes, sin embargo es común hacer estas determinaciones para conocer su valor nutrimental (Abbasi & Xinbo, 2015).
Además de los macronutrientes, el contenido de minerales como el fierro (Fe) y el manganeso (Mn) es de interés tanto por su aporte nutrimental como para el metabolismo vegetal. El Fe es un elemento esencial para el metabolismo celular como cofactor de numerosas enzimas (Wandersman & Delepelaire, 2004), además tiene diversas funciones en procesos biológicos esenciales, como transporte de oxígeno, síntesis de ácido desoxirribonucleico (ADN), fijación de nitrógeno, respiración y fotosíntesis (Greenshields, Guosheng, Feng, Selvaraj & Wei, 2007). Sin embargo, altas concentraciones de este elemento pueden producir estrés oxidativo en las plantas (Bienfait, 1988). Se considera que el contenido normal de Fe en el tejido vegetal es de 50 ppm a 300 ppm (Yi & Guerinot, 1996).
El Mn es un elemento necesario para muchos procesos enzimáticos y redox en las plantas, así como en la fotosíntesis (Memon, Aktoprakligül, Zdemur & Verti, 2001), no obstante, altas concentraciones de Mn (> 700 mg/Kg) pueden resultar tóxicas para las plantas (Markert, 1992). El Mn forma complejos estables con moléculas biológicas, cataliza la formación de la clorofila y las reacciones de óxido-reducción en los tejidos. En las plantas es un elemento poco móvil y en el suelo se encuentra en compuestos análogos a los del Fe. La disponibilidad de Fe y Mn es limitada en suelos que presentan valores altos de pH o que presentan carbonatos libres. La deficiencia de Fe y Mn en las dicotiledóneas se caracteriza, a menudo, por pequeñas manchas amarillas, o clorosis intervenal; las hojas superiores de las plantas con deficiencia se vuelven amarillentas entre nervaduras, mientras que estas permanecen verdes (Hsieh et al., 2012). Considerando que el Fe y el Mn detectados a nivel de planta son absorbidos de la rizósfera y que son transportados hasta la semilla (Schulte & Kellingg, 1999), que es la parte comestible de la planta, en este trabajo se tuvo como objetivo conocer el contenido de Fe y Mn en hojas y tallos y de macronutrientes en las semillas de chamal (Dioon edule).
MATERIALES MÉTODOS
Material vegetal
Los frutos maduros de chamal se colectaron de plantas adultas localizadas en la comunidad Vergel 2 (sitio 2, latitud N: 21º83'94.47” y longitud W: 99º38'6475”) con las del municipio de Tamasopo, San Luis Potosí, México. También se obtuvieron plantas pequeñas en las comunidades Vergel 1 (sitio 1, latitud N: 21º84'66.47” y longitud W: 99º38'99.01”), Vergel 2 (sitio 2, latitud N: 21º83'94.47” y longitud W: 99º38'6475”), Ojo de Agua (sitio 3, latitud N: 22o08'84.09” y longitud W: 99º40'68.48”) y Providencia (sitio 4, latitud N: 21º85'86.58” y longitud W: 99º39'13.56”) del mismo municipio. Las plantas colectadas se aclimataron en condiciones de vivero para la producción de semillas. Las semillas de maíz amarillo y frijol negro variedad Michigan se adquirieron en el mercado local de Ciudad Valles, San Luis Potosí, México.
Procesamiento de plantas de D. edule
Las plantas de chamal colectadas en campo se lavaron con agua potable, seguido de agua desionizada y, finalmente, con una solución 0.01 M de ácido etilendiaminotetracético (EDTA) para remover los metales adsorbidos al tejido vegetal (Carranza-Álvarez, Alonso-Castro, Alfaro de la Torre & García de la Cruz, 2008). Las plantas se separaron en hojas y tallos y se sometieron a un proceso de secado a 70 °C durante 12 h, posteriormente se pulverizaron en un molino analítico y se preservaron en oscuridad y en recipientes cerrados hasta su digestión ácida.
Digestión ácida del material vegetal
Para el análisis de Fe y Mn se realizó una digestión ácida del material vegetal (tallo, hojas y semillas de D. edule) utilizando 0.1 mL de agua regia (1HNO3: 3HCl)/mg de material vegetal durante 5 h a 70 °C. La digestión se continuó con HNO3 (0.2 mL/mg de muestra vegetal) a 60 °C durante 12 h. Posteriormente, el ácido restante se evaporó, las muestras se dejaron secar a temperatura ambiente, el residuo se resuspendió con HNO3 5% (v/v) y se completó el volumen a 25 mL (Loring & Rantala, 1992). Además, se procesaron muestras por triplicado del material vegetal de referencia Lagaroshipon major (CBR, material de referencia núm. 60) y los blancos de reactivo para determinar la exactitud de la recuperación de Fe y Mn. La tasa de recuperación para la muestra de referencia osciló entre 90% y 110%.
Cuantificación de Fe y Mn en tallo y hojas de D. edule
El contenido de Fe y Mn en el material vegetal se determinó mediante espectrofotometría de absorción atómica (EAA) acoplado a llama (Varian-SpectrAA 220 FS, Palo Alto, CA), utilizando aire-acetileno. Los límites de detección analíticos fueron de 0.03 mg/Kg y 0.01, mg/Kg para Fe y Mn, respectivamente. También se analizó una muestra de agua de concentración estándar conocida (Water for trace elements-Drinking Water Standard [TM-DWS], Nacional Investigación del Agua, Canadá) junto con las muestras para garantizar la calidad de los métodos analíticos. La tasa de recuperación de osciló entre 100% ± 15% (Carranza-Álvarez et al., 2008).
Determinación de macronutrientes
Las semillas de chamal y granos enteros crudos de maíz y frijol se pulverizaron en un molino analítico hasta que el tamaño de la partícula pasó por el tamiz de malla 110 (148 μm de apertura). Las harinas se almacenaron en un lugar fresco y seco hasta su uso. A las harinas de semillas de chamal, maíz y frijol se les realizó un análisis químico proximal de acuerdo con las técnicas descritas en Association of Official Analytical Chemist (AOAC, 2012); humedad (Método 925.09) en estufa de secado (marca Binder Mod. ED 53 UL) a 110 °C, cenizas (Método 923.03) por incineración en mufla (marca Terlab) a 550 ºC ± 1 °C, proteína cruda (Método 977.14) por el método de Kjeldalh en digestor y destilador (marca Buchi), lípidos en equipo Soxhlet (Método 920.29) y los carbohidratos se calcularon por diferencia de los otros macronutrientes (Diario Oficial de la Federación [DOF], 2010). Cada determinación se hizo por triplicado.
Contenido energético
El aporte de energía de las semillas de D. edule se calculó de acuerdo con lo descrito por Ledesma, Chávez, Pérez-Gil, Mendoza & Calvo (2010) y se reportó en KJ y en Kcal como se indica en la NOM-051-SSA-2010 (DOF, 2010): 1 g de proteína, grasa y carbohidratos, aportan 4 Kcal, 9 Kcal y 4 Kcal, respectivamente; y 4 Kcal equivalen a 17 KJ.
Análisis estadístico
Los resultados de cada determinación se realizaron por triplicado y se expresaron como medias ± desviación estándar y se analizaron por una prueba de ANOVA y la prueba de Tukey con un valor de p < 0.05 utilizando el programa Sigma Plot.
RESULTADOS
Contenido de Fe y Mn en tallo y hojas
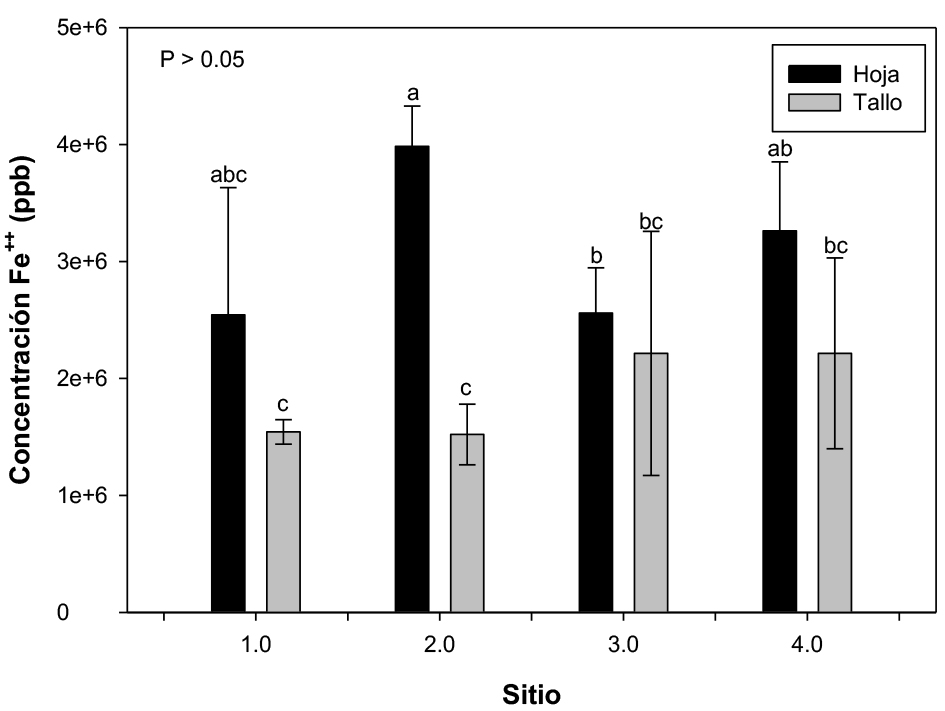
El análisis del contenido de Fe en hojas y tallos de D. edule de plantas de cuatro diferentes sitios reveló que una alta acumulación de este elemento se presenta principalmente en las hojas (figura 1). En el sitio 2 se detectó la máxima concentración a nivel de hojas (4295.1 mg/kg), seguido de la concentración en el tallo (1685.7 mg/kg). La concentración de Fe detectada en hoja de las plantas colectadas en el sitio 1 y sitio 4 fue muy similar a la detectada en el sitio 2, mientras que en el sitio 3 se detectó la menor concentración. El análisis estadístico no demostró diferencia significativas entre los sitios de colecta, pero sí entre el tejido analizado en cada sitio (figura 1).
|
|
|
 |
|
|
Figura 1. Concentración media (n = 3) de Fe en plantas de Dioon edule provenientes de cuatro sitios diferentes. Las barras de error representan el error estándar de las concentraciones. Las medias con diferentes letras muestran diferencias significativas (p < 0.05).
Fuente: Elaboración propia. |
|
Figura 1. Concentración media (n = 3) de Fe en plantas de Dioon edule provenientes de cuatro sitios diferentes. Las barras de error representan el error estándar de las concentraciones. Las medias con diferentes letras muestran diferencias significativas (p < 0.05).
Fuente: Elaboración propia. Close |
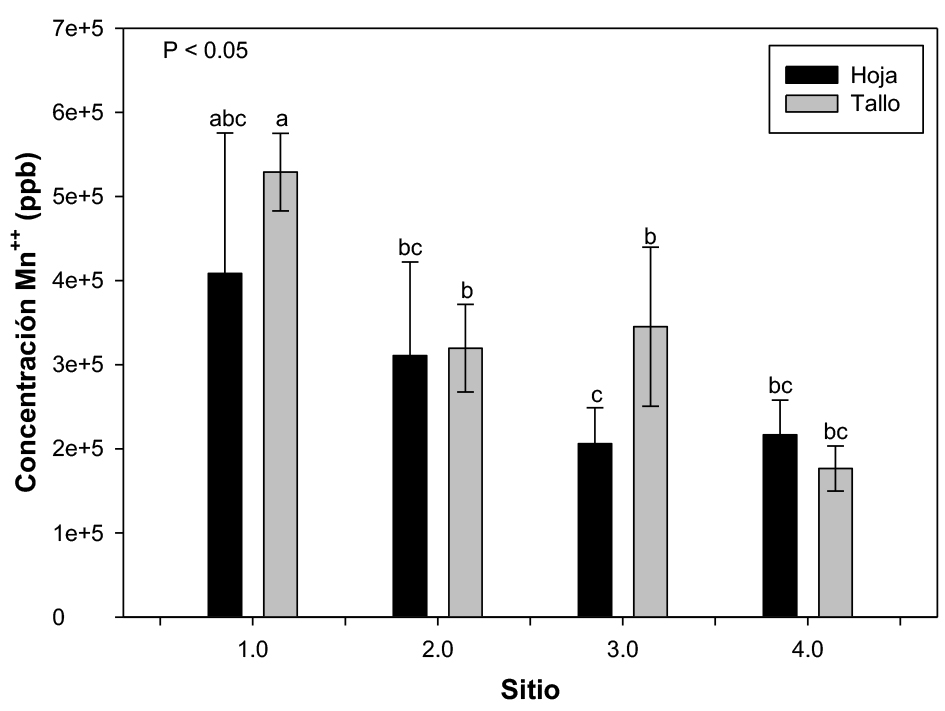
Se encontró que la acumulación de Mn tanto en tallo como en hoja es muy similar. En el sitio 1 se detectó la concentración de Mn más alta a nivel de tallo (4153.6 mg/kg). La concentración de Mn en hoja y tallo de las plantas de chamal fue mayor en las plantas colectadas en el sitio 1 > sitio 3 > sitio 2 > sitio 4 (figura 2). El análisis estadístico reveló una diferencia estadísticamente significativa (p ≤ 0.05) entre los sitios de colecta pero no entre el tipo de tejido (hoja y tallo) (figura 2).
|
|
|
 |
|
|
Figura 2. Concentración media (n = 3) de Mn en plantas Dioon edule colectadas en cuatro sitios diferentes. Las barras de error representan el error estándar de las concentraciones. Las medias con diferentes letras muestran diferencias significativas (p < 0.05).
Fuente: Elaboración propia. |
|
Figura 1. Concentración media (n = 3) de Mn en plantas Dioon edule colectadas en cuatro sitios diferentes. Las barras de error representan el error estándar de las concentraciones. Las medias con diferentes letras muestran diferencias significativas (p < 0.05).
Fuente: Elaboración propia. Close |
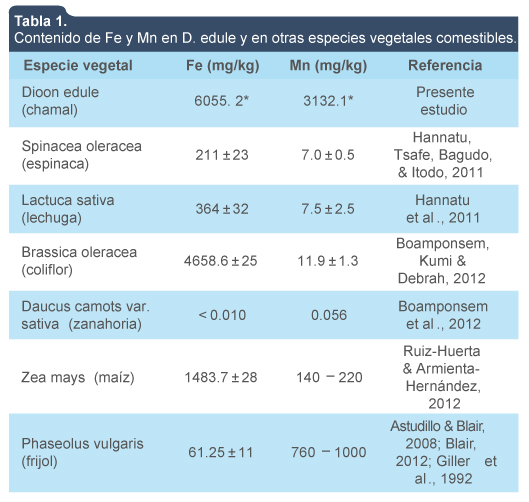
La tabla 1 muestra un análisis comparativo de la concentración total de Fe y Mn en otras especies vegetales comestibles y chamal. El análisis reveló que las plantas de chamal aportan una mayor cantidad de Fe y Mn que otras especies vegetales comestibles. De acuerdo con Sawidis, Chettri, Zachariadis & Stratis (1995), aún entre especies del mismo género pueden encontrarse diferencias en la acumulación de minerales, ya que la absorción de elementos como Fe y Mn no solo sigue los patrones fisicoquímicos de la rizosfera, sino que está regulada por las condiciones fisiológicas de las plantas y de la biota asociada a ellas para favorecer la biodisponibilidad y absorción de los elementos (Kothari, Marschner & Römheld, 1991). Es decir, las altas concentraciones de Fe y Mn detectadas en las plantas de chamal pueden atribuirse a la alta concentración de estos elementos en el suelo y a la capacidad de la planta para traslocarlos a tejidos aéreos. Se requieren los elementos en cantidades variables dependiendo de la etapa de desarrollo y diferentes condiciones ambientales y de estrés. Para ello, las plantas han desarrollado diferentes estrategias adaptables para abastecer los procesos metabólicos y minimizar los efectos nocivos de los excesos y déficits de nutrientes minerales en el medio ambiente, para así lograr una cantidad suficiente de elementos esenciales, evitar la acumulación excesiva de elementos que podría ser tóxico y para hacer frente a los efectos nocivos de elementos no esenciales. Estas estrategias incluyen mecanismos que van desde funciones estructurales en las proteínas, el equilibrio osmótico, el potencial electroquímico, señalización y activación de enzimas (Williams & Salt, 2009).
Tabla 1.
Contenido de Fe y Mn en D. edule y en otras especies vegetales comestibles.
|
|
Especie vegetal
|
Fe (mg/kg)
|
Mn (mg/kg)
|
Referencia
|
|
Dioon edule (chamal)
|
6055. 2*
|
3132.1*
|
Presente
estudio
|
|
Spinacea oleracea (espinaca)
|
211 ± 23
|
7.0 ± 0.5
|
Hannatu,
Tsafe, Bagudo,
& Itodo, 2011
|
|
Lactuca sativa (lechuga)
|
364 ± 32
|
7.5 ± 2.5
|
Hannatu
et al., 2011
|
|
Brassica oleracea (coliflor)
|
4658.6 ± 25
|
11.9 ± 1.3
|
Boamponsem,
Kumi & Debrah, 2012
|
|
Daucus camots var.sativa (zanahoria)
|
< 0.010
|
0.056
|
Boamponsem
et al., 2012
|
|
Zea mays (maíz)
|
1483.7 ± 28
|
140 – 220
|
Ruiz-Huerta
& Armienta-Hernández,
2012
|
|
Phaseolus vulgaris (frijol)
|
61.25 ± 11
|
760 – 1000
|
Astudillo & Blair,
2008; Blair,
2012; Giller
et al., 1992
|
*El promedio corresponde a las plantas completas.
Fuente: Elaboración propia.
Abrir
 |
Fuente: Elaboración propia. Close |
Contenido de macronutrientes en semillas de Chamal
En la tabla 2 se muestran los resultados del análisis químico proximal de las semillas de chamal, maíz amarillo y frijol negro variedad Michigan. Se observó que en las tres tipos de muestras el componente mayoritario fueron los carbohidratos, seguidos de las proteínas y en menor cantidad los lípidos. Las semillas de chamal tuvieron 14.67% de humedad, mientras que la del maíz y el frijol fueron de 10.8% y 6.94%, respectivamente. El contenido de cenizas para las semillas de chamal fue de 0.77%, y los del maíz y frijol fueron de 1.1% y 3.68%, respectivamente.
Tabla 2.
Resultados del análisis químico proximal de semillas de chamal (Dioon edule), maíz amarillo (Zea mays) y frijol (Phaseolus vulgaris).
|
|
Composición
próxima (%)
|
Valores medidos
|
|
D. edule (chamal)
|
Z. mays
|
P. vulgaris
|
|
Proteinas
|
20.65 ± 3.67a
|
9.42 ± 1.23b
|
21.90 ± 0.20a
|
|
Lípidos
|
3.21 ± 0.22a
|
4.74 ± 0.87b
|
4.45 ± 0.30b
|
|
Carbohidratos
|
60.69 ± 0.21a
|
74.26 ± 0.65b
|
63.01 ± 0.42c
|
|
Humedad
|
14.67 ± 0.29a
|
10.80 ± 0.31b
|
6.94 ± 0.21c
|
|
Cenizas
|
0.77 ± 0.06a
|
1.1 ± 0.17b
|
3.68 ± 0.09c
|
* Los valores son el promedio de tres réplicas ± DS.
Las letras diferentes entre columnas indican diferencia significativa (p < 0.05).
Fuente: Elaboración propia.
Abrir
 |
* Los valores son el promedio de tres réplicas ± DS.
Las letras diferentes entre columnas indican diferencia significativa (p < 0.05).
Fuente: Elaboración propia. Close |
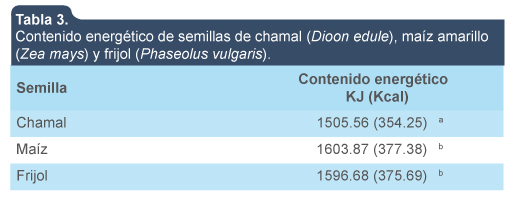
El contenido energético que aportan las semillas de chamal fue significativamente diferente (p < 0.05) al que aportan el maíz y el frijol (tabla 3).
Tabla 3.
Contenido energético de semillas de chamal (Dioon edule), maíz amarillo (Zea mays) y frijol (Phaseolus vulgaris).
|
|
Semilla
|
Contenido energético
KJ (Kcal)
|
|
Chamal
|
1505.56 (354.25)a
|
|
Maíz
|
1603.87 (377.38)b
|
|
Frijol
|
1596.68 (375.69)b
|
* Los valores son el promedio de tres réplicas ± DS.
Las letras diferentes entre columnas indican diferencia significativa (p < 0.05).
Fuente: Elaboración propia.
Abrir
 |
* Los valores son el promedio de tres réplicas ± DS.
Las letras diferentes entre columnas indican diferencia significativa (p < 0.05).
Fuente: Elaboración propia. Close |
DISCUSIÓN
La importancia del Fe en las plantas radica en sus funciones de aceptación y donación de electrones y juega un papel importante en la cadena de transporte de electrones de la fotosíntesis y respiración, por lo que es indispensable en la transferencia de energía en las especies vegetales. Además, el Fe es un constituyente de enzimas y proteínas, y al igual que en las plantas, en los seres humanos tiene importancia en el metabolismo (Ebrahimian & Bybordi, 2011).
La concentración óptima tanto de Fe y Mn en tejido vegetal se considera que debe estar en un rango de 50 mg/kg a 300 mg/kg (Jones, Wolf & Mills, 1991). Estos dos elementos tienen un importante papel en la nutrición vegetal y en la cadena alimenticia (Rehman, Aziz, Farooq, Wakeel & Rengel, 2012; Rengel, 2001; 2011).
En cuanto al contenido de Fe y Mn, las semillas de chamal aportan 98.8 y 3.1 veces más la concentración que frijol, y 4.1 y 14.2 veces más la concentración de Fe y Mn, respectivamente, que el maíz. Respecto al contenido nutrimental de las semillas, el contenido de humedad para las semillas de chamal está por debajo de estos límites máximos, lo que favorecería su conservación durante el almacenamiento, ya que limita el desarrollo de microorganismos, y además favorecería su transporte en caso de que estas semillas se comercializaran. El Codex Alimentarius (Organización Mundial de la Salud [OMS], 2007) establece en la norma CODEX STAN 153-1985 (Codex Alimentarius) un límite máximo un 15.5% de contenido de humedad en el maíz para consumo humano, y de un 15% a 19% de humedad en el frijol negro (CODEX STAN 171-1989).
Las semillas de chamal tienen mayor cantidad de proteínas (20.65 ± 3.67) que el maíz (9.42 ± 1.23), pero tienen menos lípidos (3.21 ± 0.22) y carbohidratos (60.69 ± 0.21) que el maíz y el frijol; el contenido de cenizas fue menor que el del maíz y el frijol, cuyos valores son de 1.25% – 1.40%, lo que indica un bajo contenido de minerales en la muestra. Sin embargo, las semillas de chamal aportan nutrientes y energía, similares a los del estos granos. Cárdenas, Gómez, Díaz & Camarena (2000) reportan que los cultivares de Phaseolus vulgaris presentan un bajo contenido de grasa, un alto contenido de carbohidratos y contenido medio de proteínas. El valor nutritivo de las leguminosas se debe especialmente a su aporte de proteínas (20% – 35%) y de los cereales a su aporte de carbohidratos (24% – 68%), mientras que en las semillas de chamal constituyen una mejor alternativa de proteínas para la dieta que el frijol y maíz, además de contar con un menor aporte calórico (tabla 2).
En las comunidades rurales el consumo de maíz y frijol constituye la principal fuente de energía. Los resultados de este estudio demuestran que las semillas de chamal pueden ser una alternativa para abastecer los requerimientos diarios de Fe y Mn en la dieta de las comunidades de la Huasteca Potosina. Además, los resultados obtenidos en este trabajo son similares a los de otras variedades de palmas (Astrocaryum mexicanum, Chamaedorea alternans y Chamaedorea tepejilote). Centurión-Hidalgo et al. (2009) realizaron el análisis químico proximal y del contenido de minerales que las inflorescencias de las palmas Astrocaryum mexicanum, Chamaedorea alternans y Chamaedorea tepejilote en la sierra del estado de Tabasco. Los resultados de este estudio demostraron que el mayor porcentaje de proteína cruda (25.39%) y de extracto etéreo (2.26%) se encontró en las inflorescencias de C. alternans y en C. tepejilote, así como los de cenizas (17.25%) y fibra cruda (12.16%). Con respecto a los minerales, el Fe estuvo presente en menor cantidad (13.9 mg 100 g–1) en la inflorescencia de A. mexicanum, mientras que en C. alternans y C. tepejilote los contenidos fueron de alrededor de 25 mg 100 g–1. Lo anterior indica que estas palmas son una fuente rica en proteínas, minerales y fibra dietaria, y que su consumo puede contribuir a la solución de la problemática alimentaria del país (Centurión-Hidalgo et al., 2009). El contenido de proteínas y de Fe detectado en las semillas de chamal son superiores a de las inflorescencias de C. alternans y en C. tepejilote, lo que sugiere que las semillas de chamal también pueden contribuir con los requerimientos nutricionales diarios de la dieta de las comunidades indígenas en la Huasteca Potosina.
Además, las semillas de chamal se someten a un proceso de nixtamalización similar al del maíz, por lo que el contenido nutricional puede incrmentarse (Cabrera, 1992). La nixtamalización es un proceso selectivo de las proteínas, por ejemplo, durante el cocimiento del maíz, la zeina, proteína deficiente en lisina y triptófano, disminuye sus solubilidad, mientras que la glutelina que tiene un mayor valor nutricional aumenta la solubilidad, y con ello la disponibilidad de los aminoácidos esenciales; también, después de la nixtamalización existe un aumento de 2.8 veces de lisina, de triptófano, y la relación de isoleucina a leucina se incrementa 1.8 veces (Waliszewski, Estrada & Pardio, 2003).
Lo anterior indica que el proceso de nixtamalización puede incrementar el balance de aminoácidos esenciales en las semillas de chamal, agregándoles un valor nutricional mayor a las proteínas. Además, el calcio desempeña un papel muy importante durante la nixtamalización, el tratamiento de las semillas con cal facilita la remoción del pericarpio durante la cocción y el reposo, controla la actividad microbiana, mejora el sabor, aroma, color, vida de anaquel y valor nutricional (Waliszewski et al., 2003). Es importante mencionar que en el proceso de nixtamalización al que se someten las semillas de chamal se retira la cáscara de la semilla para extraer el gametofito, lo que permite eliminar las sustancias tóxicas como el aminoácido no proteico, derivado de alanin, la β-N-metilamino-L-alanina (L-BMAA), compuesto que a concentraciones elevadas es neurotóxico para mamíferos y aves (Brener et al., 2003; Schneider, Wink, Sporer & Lounibos, 2002).
Considerando que el maíz y frijol son dos granos empleados en la mayoría de la dieta de los mexicanos (Rosado, 2001), y que la comparación en cuanto al contenido proteico de las semillas de D. edule se realizó con estas dos leguminosas, se puede considerar a las semillas de chamal como una fuente no convencional para la elaboración de alimentos similares a los que se elaboran a partir de maíz.
CONCLUSIONES
El contenido de proteínas, grasas y carbohidratos en las semillas de chamal es similar al del frijol y mayor al del maíz y son un buen aporte de Fe y Mn, por lo que pueden ser un buen sustituto del maíz durante temporadas en las que este no se cultiva, ya que para su consumo también puede someterse a un proceso de nixtamalización. Estos resultados marcan la pauta para continuar con el estudio de esta especie de palma y promover su propagación.
REFERENCIAS
Abbasi, A. M., & Xinbo Guo, X. (2015). Proximate composition, phenolic contents and in vitro antioxidant properties of Pimpinella stewartii (A wild medicinal food). Journal of Food and Nutrition Research, 35), 330-336.
Association of Official Analytical Chemist (AOAC). (2012). Official Methods of Analysis. Association of Official Analytical Chemist (16th ed.), vol. 1. Washington, D.C.: AOAC.
Astudillo, C., & Blair, M. W. (2008). Evaluación del contenido de hierro y zinc en semilla y su respuesta al nivel de fósforo en variedades de fríjol colombianas. Agronomía Colombiana, 26(3), 471-476.
Bienfait, H. F. (1988). Mechanisms in Fe-efficiency reactions of higher plants. ournal of Plant Nutrition, 1(11), 605-629.
Blair, M. W. (2012). Use of the advanced backcross-QTL method to transfer seed mineral accumulation nutrition traits from wild to Andean cultivated common beans. Theoretical and Applied Genetics, 125(5), 1015-1031.
Boamponsem, G. A., Kumi, M., & Debrah, I. (2012). Heavy Metals Accumulation In Cabbage, Lettuce And Carrot Irrigated With Wastewater From Nagodi Mining Site In Ghana. International Journal Of Scientific & Technology Research, 1(11), 124-129.
Brener, E. D., Stevenson, D. W., & Twigg, R. W. (2003). Cycads: evolutionary innovations and the role of plant-derived neurotoxins. Trends in Plant Science, 8(9), 446-452.
Cabrera, F. (1992). Diccionario de aztequismos. México: Colofon.
Cárdenas, Q. H., Gómez, B. C., Díaz, N. J., & Camarena, N. F. (2000). Evaluación de la calidad de la proteína de cuatro variedades mejoradas de frijol. Revista Cubana Alimentaria Nutricional, 14(1), 22-27.
Carranza-Álvarez, C., Alonso-Castro, A. J., Alfaro de la Torre, M. C., & García de la Cruz, R. F. (2008). Accumulation and distribution of heavy metals in Scirpus americanus and Typha latifolia from an artificial lagoon in San Luis Potosí, Mexico. Water Air Soil Pollut, 188(1), 297-309.
Centurión-Hidalgo, D., Alor-Chávez, M. J., Espinosa-Moreno, J., Gómez-García, E., Solano, M. L., & Poot-Matu, J. E. (2009). Contenido nutricional de inflorescencias de palmas en la sierra del estado de Tabasco. Universidad y Ciencia, 25(3), 193-199.
Diario Oficial de la Federación (DOF). (2010).Norma Oficial Mexicana NOM-051-SCFI/SSA1-2010. Especificaciones generales de etiquetado para alimentos y bebidas no alcohólicas preenvasados-Información comercial y sanitaria. Recuperado el 25 de enero de 2017 de http://dof.gob.mx/nota_detalle.php?codigo=5137518&fecha=05/04/2010.
Ebrahimian, E., & Bybordi, A. (2011). Effect of iron foliar fertilization on growth, seed and oil yield of sunflower grown under different irrigation regimes. Middle East Journal of Scientific Research 9(5), 621-627.
Giller, K. E., Amijee, F., Brodrick, S. J., McGrath, S. P., Mushi, C., Edje, O. T., & Smithson, J. B. (1992). Toxic concentrations of iron and manganese in leaves of Phaseolus vulgaris L. growing on freely‐drained soils of pH 6.5 in Northern Tanzania. Communications in Soil Science and Plant Analysis, 23(15), 1663-1669.
Greenshields L. D., Guosheng, L., Feng, J., Selvaraj, G., & Wei, Y. (2007). The siderophore biosynthetic gene SID1, but not the ferroxidase gene FET3, is required for full Fusarium graminearum virulence. Molecular Plant Pathology, 8(4), 411-421.
Hannatu, A. S., Tsafe, A. I., Bagudo, B. U., & Itodo, B. U. (2011). Toxic Metals Uptake by Spinach (Spinacea oleracea) and Lettuce (Lactuca sativa) Cultivated in Sokoto: A Comparative Study. Pakistan Journal of Nutrition, 10(6), 572-576.
Hsieh, S. I., Castruita, M., Malasarn, D., Urzica, E., Erde, J., Page, M. D., Yamasaki, H., Casero, D., Matteo Pellegrini, M., Merchant, S. S., & Loo, J. A. (2012). The Proteome of Copper, Iron, Zinc, and Manganese Micronutrient Deficiency in Chlamydomonas reinhardtii. Molecular Cell Proteomics, 12(1), 65-86.
Jones, J. B., Wolf, B., & Mills, H. A. (1991). Plant analysis handbook: a practical sampling, preparation, analysis, and interpretation guide. Athens, GA, USA: Macro-Micro Publ.
Kothari, S. K., Marschner, H., & Römheld, V. (1991). Effect of a vesicular-arbuscular mycorrhizal fungus and rhizosphere microorganisms on manganese reduction in the rhizosphere and manganese concentrations in maize (Zea mays L.). New Phytol, 117(4), 649-655.
Ledesma Solano, J. A., Chávez Villasana, A., Pérez-Gil Romo, F., Mendoza Martínez, E., & Calvo Carrillo, C. (2010). Composición de alimentos. Miriam Muñoz de Chávez. Valor nutritivo de los alimentos de mayor consumo(2da. ed.), México: McGrawHill.
Loring, D. H., & Rantala, R. T. T. (1992). Manual for the geochemical analyses of marine sediments and suspended particulate matter. Earth-Science Reviews, 32(4), 235-283.
Market, B. (1992). Presence and significance of naturally occurring chemical elements of periodic system in the plant organism and consequences for future investigation on inorganic environmental chemistry in ecosistems. Vegetation, 103(1), 1-30.
Memon, A. R., Aktoprakligül, D., Zdemur, A., & Verti, A. (2001). Heavy metal accumulation and detoxification mechanisms in plants. Turkish Journal of Botany, 25(3), 111-21.
Organización Mundial de la Salud (OMS). (2007). Codex Alimentarius. Roma: Organización de las Naciones Unidas para la Agricultura y la Alimentación. Recuperado el 25 de enero de 2017 de http://files.foodmate.com/2013/files_1113.html
Rehman, H., Aziz, T., Farooq, M., Wakeel, A., & Rengel, Z. (2012). Zinc nutrition in rice production systems: a review. Plant and Soil, 361(1), 203-226.
Rengel, Z. (2001). Genotypic differences in micronutrient use efficiency in crops. Communication in Soil Science and Plant Analysis, 32(7-8), 1163-1186.
Rengel, Z. (2011). Soil pH, soil health and climate change. In B. P. Singh, A. L. Cowie & K. Y. Chan (ed.), Soil Health and Climate Change (pp. 68-95). Berlin Heidelberg: Springer.
Rosado, L. J. (2001). Dietary fiber in Mexico: Recomendations and actual consumption patterns. In Susan Sungsoo Cho & Mark L. Dreher (Ed.), Handbook of Dietary Fiber. New York: Marcel Dekker Inc.
Ruiz-Huerta, E. A., & Armienta-Hernández, M. A. (2012). Acumulación de arsénico y metales pesados en maíz en suelos cercanos a jales o residuos mineros. Revista Internacional de Contaminación Ambiental, 28(2), 103-117.
Sawidis, T., Chettri, M. K., Zachariadis, G. A., & Stratis, J. A. (1995). Heavy metals in aquatic plants and sediments from water systems in Macedonia, Greece. Ecotoxicology and Environmental Safety, 32(1), 73-80.
Schneider, D., Wink, M., Sporer, F., & Lounibos, P. (2002). Cycads: their evolution, toxins, herbivores and insect pollinators. Naturwissenschaften, 89(7), 281-294.
Schulte, E. E., & Kelling, K.A. (1999). Soil and applied manganese. En Understanding Plant Nutrients, A2526. USA: Cooperative Extension Publications, University of Wisconsin-Extension.
Waliszeski, K.N., Estrada, Y., & Pardio, V. (2003). Recovery of lysine and tryptophan from fortified nixtamalizedconrflour and tortillas. International Journal of Food Science and Technology, 3838(1), 73-75.
Wandersman, C., & Delepelaire, P. (2004). Bacterial iron sources: from siderophores to hemophores. Annual Review of Microbiology , 58, 611-647.
Williams, L., & Salt, D. E. (2009). The plant ionome coming into focus. Current Opinion in Plant Biology, 12(3), 247-249. Recuperado de http://doi.org/10.1016/j.pbi.2009.05.009. http://eprints.soton.ac.uk/142463/
Yáñez Espinoza, L. (2006). Las cycadas: biología y conservación en México. Estado de México: Universidad Autónoma de Chapingo.
Yi, Y., & Guerinot, M. L. (1996). Genetic evidence that induction of root Fe (III) chelate reductase activity is necessary for iron uptake under iron deficiency. Plant Journal, 10(5), 835-844.